
Application Note
Automated Monitoring of Development and Analysis of iPSC-derived 3D Cerebral Organoids
- Improve 3D biology workflows with easy-to-use deep learning-based analysis tools
- Develop methods that better represent cell self-organization that is consistent with in vivo development
- Automate culturing and monitoring of cerebral organoids
Angeline Lim | Sr. Applications Scientist | Molecular Devices
Oksana Sirenko | Sr. Manager of Assay Development | Molecular Devices
Introduction
The human brain is highly complex which makes it challenging to study both in vitro and in-model organisms. Cultured neurons do not sufficiently recapitulate the three-dimensional (3D) architecture, complexity, and microenvironment of the brain. And while the use of model organisms has helped define some elements of conserved developmental pathways, suitable models for studying certain diseases—such age-related neurological disorders —are lacking.
However, the use of induced pluripotent stem cell (iPSC)-derived brain organoids is a promising model – it recapitulates aspects of human neurodevelopment, self-organizes into discrete brain regions, and their gene expression pattern is similar to in vivo data.1,2 Previous efforts have led to the development of several protocols to culture brain organoid and techniques that enabled the study of brain development and the diseases associated with it—most notably, the ability to culture cerebral organoids allowed for the study of human cortical development and its related diseases. Further work is required before cerebral organoids can be used at scale for functional genomics studies, drug discovery, and the study of compound-induced toxicity effects.
Here, we describe a method for semi-automated culturing and monitoring of cerebral organoids, as well as the testing of functional neuronal activity by means of recording Ca2+ oscillations. Cerebral organoids were developed from iPSC using previously published methods.2,3 Our method was adapted for a partially automated workflow and culture monitoring using the ImageXpress® Micro Confocal High-Content Imaging System. We monitored the size and morphology of developing brain microtissues in transmitted light over 20 weeks of development. Deep learning-based image analysis from IN Carta® Image Analysis Software was used for defining the size and shape of the tissues. Selected microtissues were analyzed during the different phases of development using confocal imaging by expression of Sox2, TuJ1, and GAFP markers. For detection of functional activity, calcium oscillations from selected clusters of the microtissues were recorded by imaging. Oscillations were observed after day 50 in culture and were sporadic through the volume of micro-tissue.
Materials and methods
Cell culture
Human iPSC cells adapted to feeder-free conditions (SC102A-1, System Biosciences) were thawed and cultured in Complete mTeSR™ Plus culture medium (STEMCELL Technologies) in Matrigel-coated plates (cat. #354277, Corning). Media was changed every day except once per week when a double volume of media was added to skip changing media on one day. Cells were passaged every 4-5 days using the enzyme-free reagent ReLeSR™ (STEMCELL Technologies) at a 1:6–1:10 split ratio.
Cerebral Organoid differentiation
All reagents used to generate cerebral organoids were from the STEMdiffTM Cerebral Organoid Kit (STEMCELL Technologies #08570). The kit has been optimized for cerebral organoid formation based on Lancaster et al.1 Briefly, iPSC cells were seeded at 9000 cells/well in the embryoid body (EB) seeding media in a 96-well ultra-low attachment plate (Corning). The media was replaced with EB formation media. On day 5, EBs were imaged to ensure they were ready for neural induction. EBs were transferred to a 24-well plate containing the induction medium. On day 7, EBs were transferred into Matrigel (Corning) droplets. About 6–8 droplets were placed in each well of a 6-well plate containing Maturation medium. The plate containing organoids were allowed to mature in a 37°C incubator on an orbital shaker. Fresh media was replaced every 3–4 days.
Calcium flux assay
Calcium flux was assessed using the FLIPR® Calcium 6 Assay Kit (Molecular Devices) according to the manufacturer protocol. Starting from day 17, one cerebral organoid was loaded with dye for 2 hrs and then imaged on the ImageXpress Micro Confocal.
Immunofluorescence
Whole organoids were fixed in 4% PFA overnight at 4°C, followed by a quick wash with PBS. Permeabilization was done with 0.5% triton-X in PBS. Organoids were incubated with the following antibodies and stains for 48 hours: Hoechst (33 µM, Invitrogen cat. #33342), Alexa Fluor 555 mouse anti-ß-tubulin (1:100, Becton Dickinson cat. #560339), Alexa Fluor 647 mouse anti-Sox2 (1:100, Becton Dickinson cat. #562139), Alexa Fluor Plus 750 Phalloidin (ThermoFisher Scientific cat. #A30105).
Image acquisition and analysis
All images were acquired on the ImageXpress Micro Confocal system (Molecular Devices) using MetaXpress® High-Content Image Acquisition and Analysis Software. IN Carta™ Image Analysis Software was used for all analysis. The IN Carta SINAP module was used to carry out segmentation of images. Each model was trained and verified before being used in the analysis protocol. For calcium imaging, images were acquired with camera binning set to 2. To increase acquisition speed, the 50 µm confocal slit module was used.
Results
Culture and differentiation of iPSC-derived cerebral organoids
Cerebral organoids are cultured according to the protocol adapted from Lancaster et al.3 In this method, the organoids are grown in a culture media that promotes self-organization and patterning. Embryoid bodies are further differentiated in an extracellular matrix (Matrigel) to improve polarization of the neuroepithelia as well as support the growth of large neuroepithelia buds. These buds extend from the EBs and contain cavities similar to brain ventricles. Figure 1 illustrates the main steps used to generate iPSC-derived cerebral organoids.
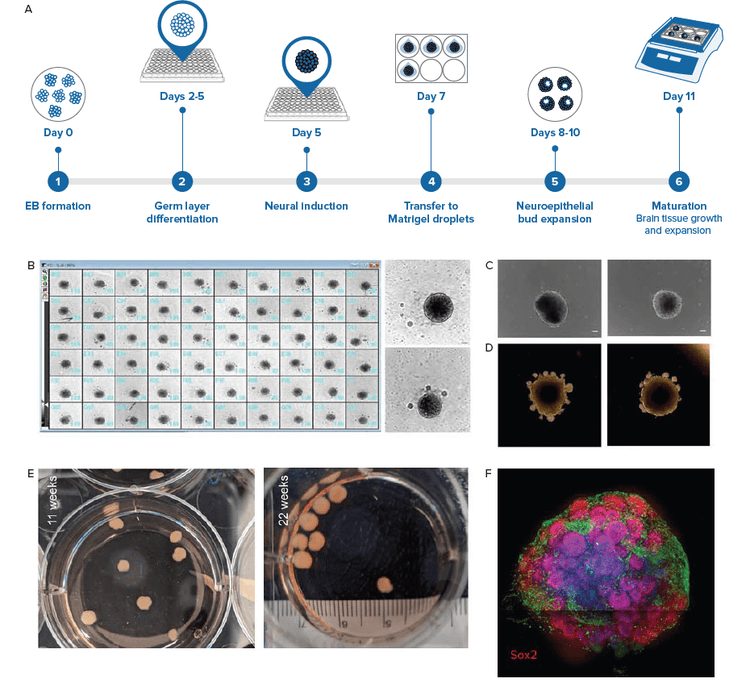
Figure 1. iPSC-derived cerebral organoids generated using the intrinsic patterning approach. iPSC cells were used to create EBs. A) Main steps used to generate iPSC derived cerebral organoids. The protocol used is based on Lancaster & Knoblich, 2014 using media from STEMCELL Technologies. B) By day 5, majority of EBs were at least 400 μm in diameter. The EBs show smooth outlines and regions near the surface were optically brighter (in brightfield microscopy). Example of EBs from plate overview enlarged on right. Scale bar = 100 μm. C) Images of EBs after they were transferred to neural induction medium. Optically translucent edges suggest formation of neuroepithelia. Scale bar = 100μm. D) From day 8–10, organoids were embedded in Matrigel to support expansion of neuroepithelia. Note the formation of numerous epithelial buds. E) Organoids were transferred to an orbital shaker and allowed to mature. Digital camera imaged cerebral organoids during maturation. F) Organoids were fixed and stained with Hoechst (nuclei, blue), TUJ1 (neuron, green), and SOX2 (radial glia, in red). Dark line across the image is a result of image tiling.
Monitor developing cerebral organoids using AI-based analysis tools
To monitor the quality of developing organoids, we used AI-based segmentation to analyze images acquired with brightfield imaging. Growth of EBs and organoids can be monitored by measuring their diameters over time. One of the advantages of using deep learning-based methods to identify EBs includes more robust segmentation. The model is trained to exclude most debris, dust particles, and imaging artifacts.
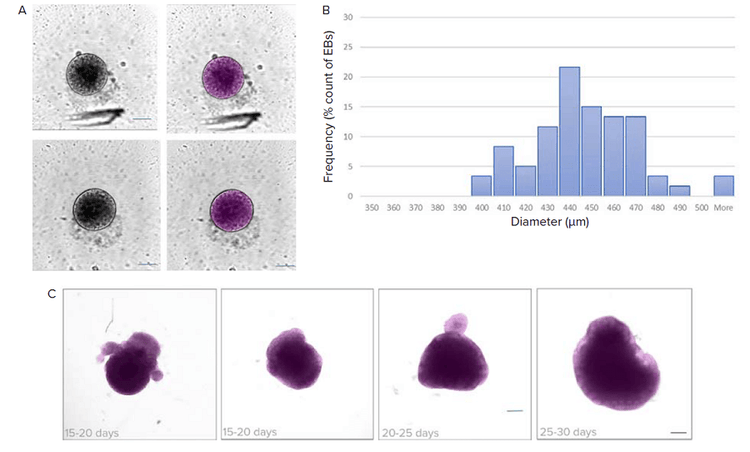
Figure 2. Analysis of brightfield images using deep learning-based segmentation (SINAP). A) EBs were monitored for their size before proceeding to the neural induction step. Shown are examples of images and their corresponding segmentation masks (in magenta) using the SINAP tool in IN Carta software. B) Histogram shows distribution of EB diameter. Binned size = 10 μm. C) Maturation of organoids can be monitored using brightfield imaging and analyzed using IN Carta SINAP tool. Custom deep-learning model was trained and then applied to the dataset. Examples of organoids with segmentation mask (magenta) shown. Note that the SINAP model allows for segmentation of organoids with different shapes and sizes. Scale bar = 100 μm.
To determine the cell types that make up the organoid, selected organoids were fixed and immunostained with SOX2 (radial glial), TUJ1 (neurons) and Hoechst (nuclei) at various time points. Cerebral organoids show organization reminiscent of a developing brain. Radial glial cells were found surrounding ventricles in the organoids. Older organoids (week 8) had larger but fewer ventricles compared to less mature (week 4) organoids. (Figure 3)
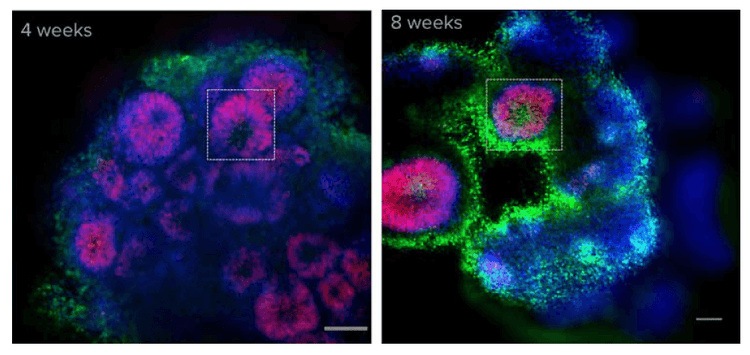
Figure 3. Cerebral organoids show organization reminiscent of a developing brain. After calcium imaging, organoids were fixed and stained with Hoechst (blue) and SOX2 (radial glia, in red). Shown here is one optical section. Ventricles (boxed) can be observed in both 4- and 8-week old organoids. SOX2 staining indicates that radial glial cells are localized near the ventricles, in a pattern similar to the cellular organization in the developing cortexcortex.3 Scale bar = 100 μm.
Calcium imaging of whole brain organoids
Calcium imaging can be used as a functional assay to determine neuronal activity.4 Starting from week 4, we observed calcium activity in cerebral organoid loaded with FLIPR Calcium 6 dye. (Figure 4). Frequency of calcium activity was low, suggesting that the neurons in the organoid at that time were still immature. By week 13, the calcium activity appeared more synchronous, suggesting that neurons are interconnected in a functional network (Figure 5).
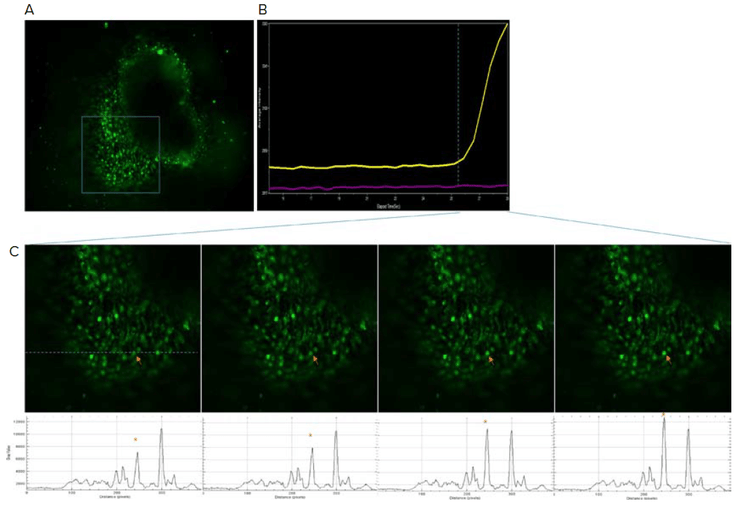
Figure 4. Calcium activity present as early as week four. A) Organoid was loaded with FLIPR Calcium 6 dye and calcium flux activity was monitored on the ImageXpress Micro Confocal system. B) Calcium intensity plot over time shown on the right. The yellow trace corresponds to the region indicated by the arrow in (C). Purple trace represents a neighboring region. C) Close up view of boxed region from (A). Intensity profile along the dotted line shown at the bottom. Arrow corresponds to the asterisk peak in the intensity profile.
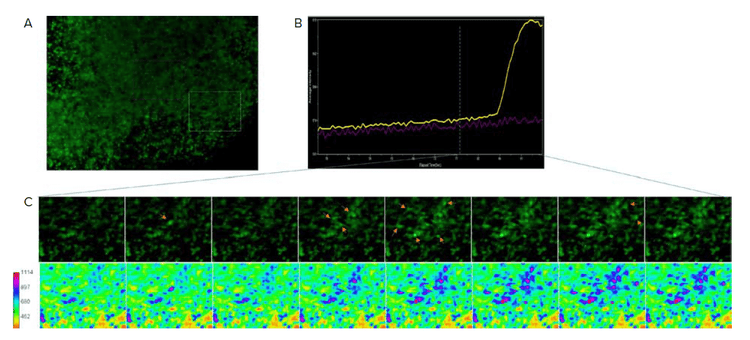
Figure 5. Synchronous calcium activity by week 13. A) Image shows an optical section of calcium 6 loaded organoid. B) Calcium intensity from boxed region represented as average intensity over time. C) Close up view of box region from (A) shown over time. Arrows indicate elevated intensity. Bottom images represent calcium intensity as a heat map. Note the intensity spreading from the initial spike suggesting presence of neuronal network.
To explore the feasibility of using these cerebral organoids in toxicity studies, organoids were incubated with compounds such as 4-Aminopyridine (neuro-stimulant) and muscimol (neuro-depressant). In some cases, 4-AP led to activation of calcium activity (Figure 6).
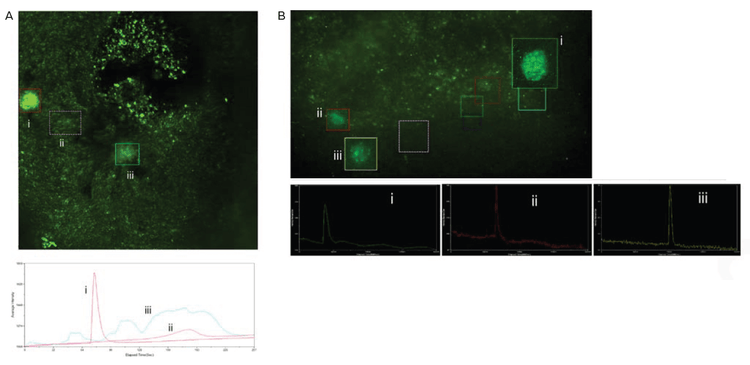
Figure 6. Calcium activity triggered by neuro-stimulants. A) Organoid were incubated with 100nM of 4-AP for 30mins and then imaged. Numerous calcium signaling events were observed. Boxed regions indicate areas where calcium spikes were observed. Their corresponding intensity profiles are shown below. B) Organoids treated with muscimol. Shown here is an example of recovery of calcium activity after muscimol was removed. Inhibitory effect of GABA was inconclusive due to the transient/inconsistent activity of organoids. Calcium signaling events shown in boxed regions along with their intensity profiles.
Conclusions
- We demonstrate here the workflow and tools used to automate imaging and monitor the development of cerebral organoids.
- The presence of calcium flux and the organization of the cells within the organoid is consistent with in vivo cortical development.
- The presence of calcium flux and the organization of the cells within the organoid is consistent with in vivo cortical development.
References
- Gordon, A., Yoon, SJ., Tran, S.S. et al. Long-term maturation of human cortical organoids matches key early postnatal transitions. Nat Neurosci 24, 331–342 (2021).
- Lancaster, M., Renner, M., Martin, CA. et al. Cerebral organoids model human brain development and microcephaly. Nature 501, 373–379 (2013).
- Lancaster MA, Knoblich JA. Generation of cerebral organoids from human pluripotent stem cells. Nat Protoc. Oct 9(10), 2329–40 (2014)
- Grienberger C, Konnerth A. Imaging calcium in neurons. Neuron 73, 862–885 (2012)